DUAL-PAM-100
Version:
P700 & Chlorophyll Fluorescence Measuring System
M. Onishi, R. Furuya, C. Miyake (Kobe), G. Schansker (Walz)
Simultaneous measurement of fast fluorescence and P700 induction at high light intensities (O-I1-I2-P-type measurement) is a powerful tool to probe lesions in the electron transport chain. Mineral deficiency is an example that can illustrate this.
The O-I1-I2-P or OJIP transient reflects the reduction of the electron transport chain, with the O-I1 rise paralleling the reduction of the acceptor side of PS II, the I1-I2 rise paralleling the reduction of the PQ pool, and the I2-P rise paralleling the re-reduction of plastocyanin and P700. The redox changes in P700 measured simultaneously indicate an initial oxidation of P700 and a subsequent re-reduction phase. This is illustrated in the next figure.
Mineral deficiencies affect these dynamics in different ways, and two of such effects are described here. One is the reaction induced by deficiencies of P (phosphorus) and Cu (copper), and the other by deficiencies of Zn (zinc), Ca (calcium), K (potassium), and N (nitrogen).
Copper is a cofactor for plastocyanin, a small protein that transports electrons through the lumen between the cellular b6f complex and photosystem (PS)I. Copper deficiency decreases plastocyanin content and slows electron transport between the cellular b6f complex and PS I. Phosphorus in the form of phosphate, on the other hand, is required for ATP synthesis by ATP synthase; P deficiency decreases phosphate content and inhibits ATP synthase activity. Protons pass from the lumen to the stroma through ATP synthase as part of its catalytic mechanism; inhibition of ATP synthase promotes acidification of the lumen, resulting in a “steady state” of the lumen pH. As a result, plastoquinol reoxidation by the cyt b6f complex slows down electron transfer to P700.
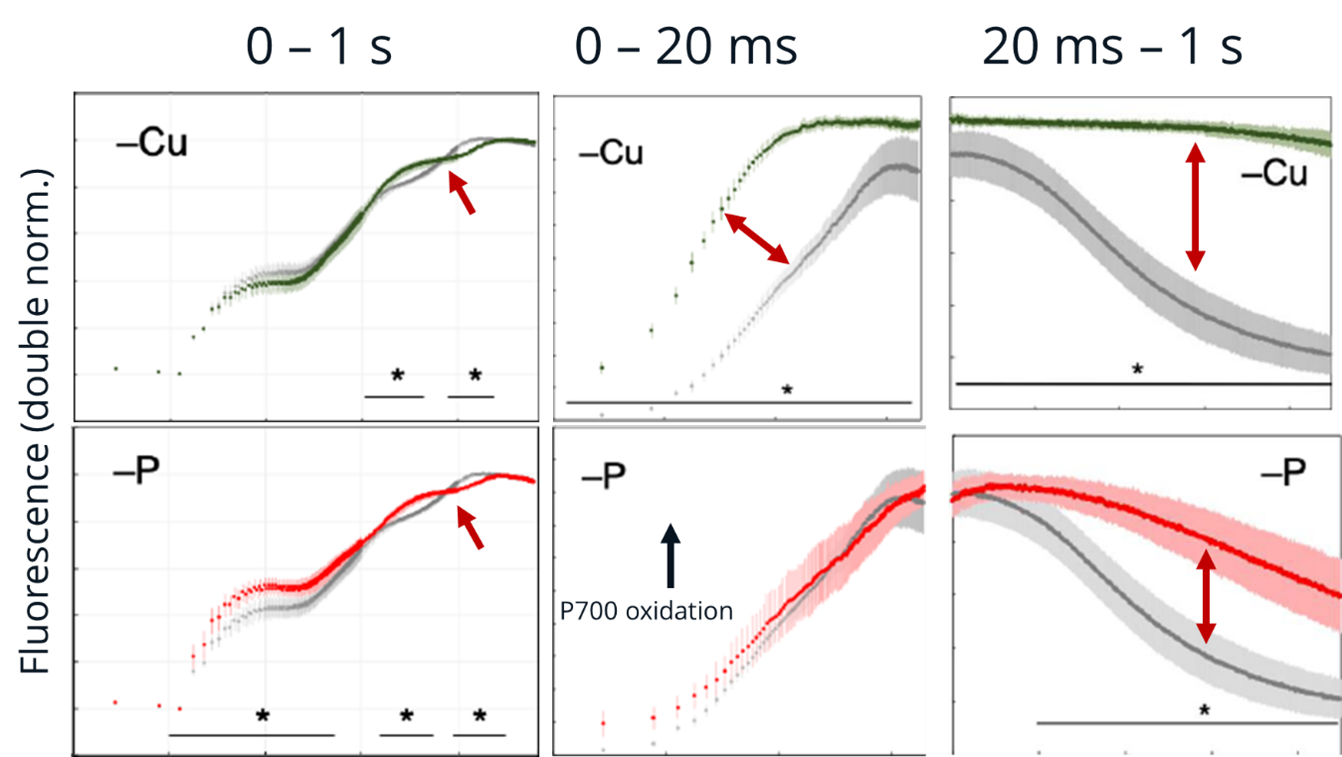
The figure shows the effect of these two deficiencies on fluorescence and P700 kinetics. In wheat, the amplitude of the I2-P phase is approximately halved and the P700+ redox kinetics are strongly slowed down in both cases. Focusing on the initial P700 oxidation phase, the difference is that P deficiency does not affect these kinetics (low lumen pH requires more time to establish), whereas low plastocyanin content (confirmed by DUAL-KLAS-NIR measurements) results in much faster P700 oxidation kinetics.
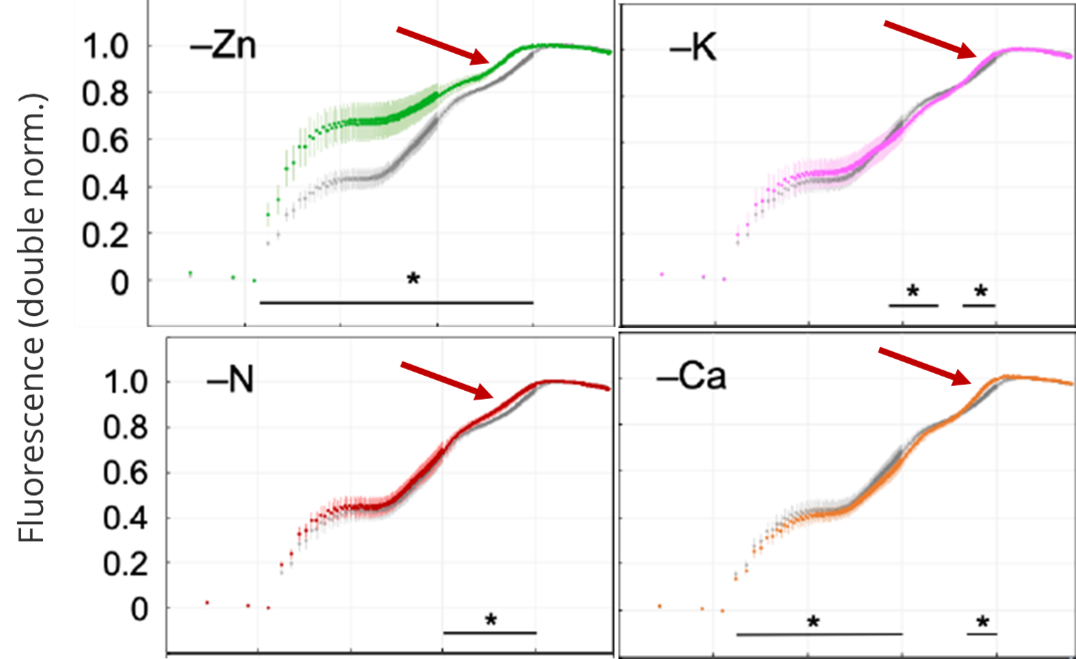
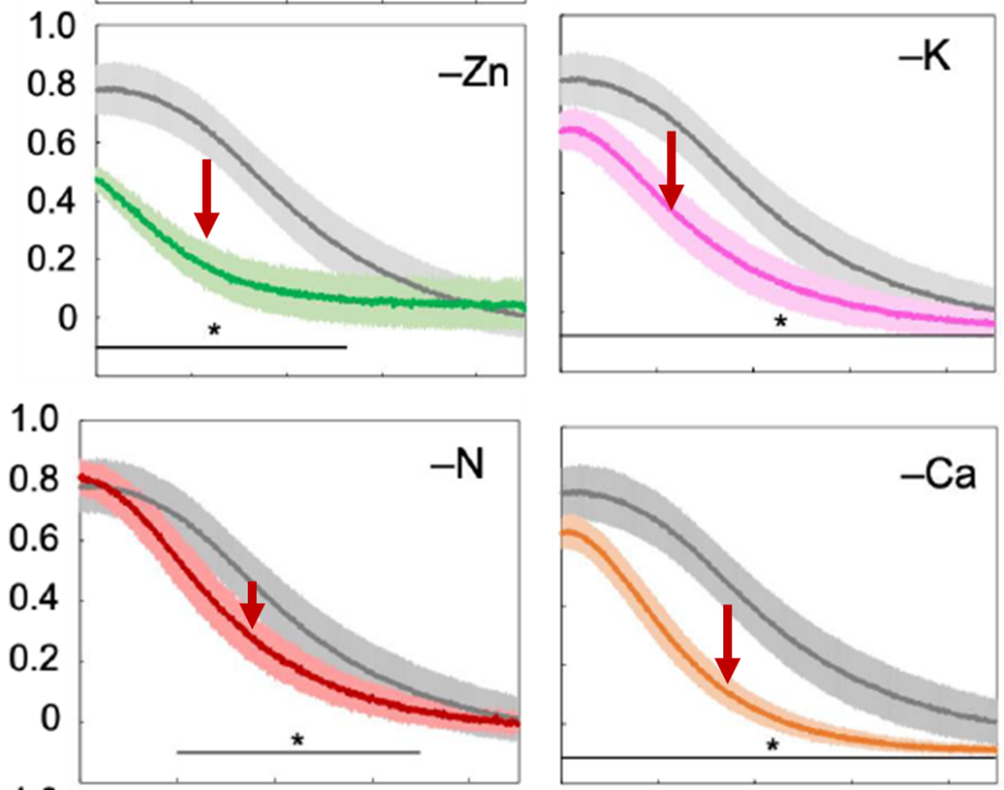
According to the literature, Zn, Ca, K and N play a role in the detoxification of oxygen radicals on the acceptor side of PS I. Deficiency is thought to cause oxygen radical-mediated damage to the acceptor side of PS I. If we look again at the dynamics of fluorescence and P700, what do we see?
The most striking effect observed in both sunflower and wheat leaves was a much faster P700+ re-reduction phase. In wheat, this was associated with a somewhat faster I2-P rise, but was not as clearly observed in sunflower leaves (Schansker et al. 2022). This can be explained by assuming that damage on the PS I acceptor side also implies a reduction in the number of electron acceptors. This would mean that fewer electrons need to flow through the electron transport chain to reduce P700+, meaning that in such leaves FM and full re-reduction of P700+ are reached earlier.
There are also a few mineral deficiencies that do not directly affect the electron transport chain but act further downstream and whose effects can be detected by feedback inhibition. Two examples (molybdenum (Mo) and boron (B) deficiencies) are shown below; the leaves were illuminated for 10 minutes, and quenching analysis was applied. Molybdenum and boron deficiencies showed higher Y(NA)-values (green) after 5 and 7.5 min of illumination, which can be interpreted as inhibition/limitation on the PS I acceptor side.
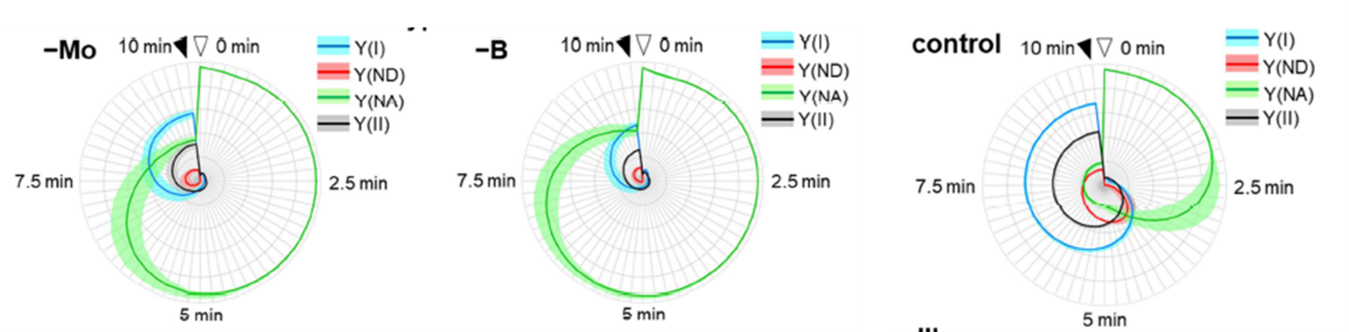
Effects induced by other deficiencies than Mo and B are observed by this type of Quenching Analysis measuring both Chl fluorescence and P700. The analysis of the visualized deficiency profile of each element will require further research. It shows us the world of the relationships between photosynthesis and minerals. It is expected that the solving of all of these relationships will create a research field by itself.
Literature
Ohnishi M, Furutani R, Sohtome T, Suzuki T, Wada S, Tanaka S, Ifuku K, Ueno D, Miyake C (2021) Photosynthetic parameters show a specific response to essential mineral deficiency. Antioxidants 10: 996; https://doi.org/10.3390/antiox10070996
Schansker G, Ohnishi M, Furutani R, Miyake C (2022) Photosynthetic parameters respond specifically to essential mineral deficiencies. Front Plant Sci13: 894607; https://doi.org/10.3389/fpls.2022.894607
Kuroki S, Ohnishi M, Furutani R, Tsuru K, Miyake C (2024) Nondestructive diagnosis of mineral deficiencies in wheat leaves by one- or two-shot saturation pulse measurement of chlorophyll fluorescence and P700+ absorbance with machine learning. Smart Agriculture Technology 9: 100586; https://doi.org/10.1016/j.atech.2024.100586
The 90 Degree Measuring Head Holder DUAL-H90 and special optical filters were employed to measure PAM fluorescence simultaneously in the spectral range below 700 nm (sw) and above 700 nm (lw). The experimental setup showed that sw fluorescence was more variable, resulting in higher PSII photochemical yields compared to lw fluorescence. The apparently higher PSII photochemical yields can be explained by low levels of constant photosystem I fluorescence in the sw spectral window. In the lw range, PSII photochemical yields are underestimated because of higher photosystem I background fluorescence.
PAN (2009) 2: 1 - 13 – NADPH Determination
New NADPH/9-AA module for the DUAL-PAM-100: Description, operation and examples of application.
By Ulrich Schreiber and Christof Klughammer.
The NADPH/9-AA module is a new accessory of the DUAL-PAM-100 system which excites fluorescence at 365 nm and detects fluorescence in the 420 to 580 nm range. The new module permits measuring of light-induced changes of NADPH fluorescence in suspensions of isolated chloroplasts, algae and cyanobacteria. Technical features of the module are outlined and examples of application are introduced. Also simultaneous measurements of chlorophyll (Chl) and NADPH fluorescence are presented. It is show that saturation pulses can serve for estimating the extent of NADP reduction in the steady state.
PAN (2008) 1: 27 - 35 – PSII Yield Parameters
Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method.
By Christof Klughammer and Ulrich Schreiber.
The fate of excitation energy in PS II is comprehensively described by the complementary quantum yields Y(II) + Y(NPQ) + Y(NO) = 1. It is shown that the simple expressions for Y(NO) and Y(NPQ) proposed by Genty et al. (1996), which do not contain Fo', are fully equivalent to the much more complex expressions of Kramer et al. (2004) and are valid for both lake and puddle models. The practical meaning of the complementary quantum yields is discussed.
PAN (2008) 1: 21 - 24 – Chl b Mutant
Monitoring the effects of reduced PS II antenna size on quantum yields of photosystems I and II using the DUAL-PAM-100 measuring system.
By Erhard Pfündel, Christof Klughammer and Ulrich Schreiber.
The DUAL-PAM-100 is employed to analyze Chl b-less barley leaves (Hordeum vulgare cv. Donaria mutant chlorina-f2 2800) and the corresponding wild-type leaves. The results show that the small PS II antenna size in the mutant affects both, PS I and PS II photochemistry.
PAN (2008) 1: 15 - 18 – Heat Stress
Non-photochemical fluorescence quenching and quantum yields in PS II and PS I: Analysis of heat-induced limitations using Maxi-Imaging-PAM and DUAL-PAM-100.
By Ulrich Schreiber and Christof Klughammer.
In this article the large potential of combined measurements with the Maxi version of the Imaging-PAM and the DUAL-PAM-100 are demonstrated.
PAN (2008) 1: 11 - 14 – PSI Yield Parameters
Saturation Pulse method for assessment of energy conversion in PS I.
By Christof Klughammer and Ulrich Schreiber.
The paper summarizes the theoretical background of the saturating pulse method which is utilized by the DUAL-PAM-100 instrument to determine the quantum yields of photochemical energy conversion and nonphotochemical energy dissipation in PS I.
PAN (2008) 1: 1 - 10 – Membrane Potential
New accessory for the DUAL-PAM-100: The P515/535 module and examples of its application.
By Ulrich Schreiber and Christof Klughammer.
The technical features of the P515/535 module are outlined and some typical examples of application are presented.The device provides information on membrane potential, membrane energization ("scattering"), and proton gradient, as well as on proton and electron fluxes.